
The Fifth Kingdom - Chapter 3
B
Kingdom Eumycota
Introduction
Traditional System:
Phylum 2 - Zygomycota -- conjugating Fungi
Class
Zygomycetes
Orders Mucorales,
Entomophthorales,
Zoopagales,
Kickxellales
Class
Trichomycetes
Orders Harpellales,
Amoebidiales, Asellariales, Eccrinales
Phylum 3 - Glomeromycota --
arbuscular
mycorrhizal fungi
Class
Glomeromycetes
New Phylogenetic System (more complex)
Phylum 4
Zygomycota
Subphylum Mucoromycotina
Order
Mucorales
Order
Endogonales
Order
Mortierellales
Groups currently of uncertain position
Subphylum
Entomophthoromycotina
Order
Entomophthorales
Subphylum
Zoopagomycotina
Order
Zoopagales
Subphylum
Kickxellomycotina
Order
Kickxellales
Order
Dimargaritales
Order
Harpellales
Order
Asellariales
Phylum 5 Glomeromycota
Class
Glomeromycetes
Phylum (?) Microsporidia
(e.g. Nosema,
intracellular parasites, mainly of insects, formerly considered protozoa,
but now regarded as extremely reduced fungi) see Corradi & Keeling
2009.
Phylum 4 Zygomycota
-- Conjugating Fungi
The second traditional eumycotan phylum
was the Zygomycota. This phylum contained two classes, class Zygomycetes
and class Trichomycetes. Since most
Trichomycetes are parasites or commensals inside the guts of living arthropods, they are
only a footnote, albeit a fascinating one. Basic
features Although this assemblage contains only about
1% of the known species of fungi, its members are distinctive, and some of them are
common, successful, fast-growing, primary colonizers of substrates containing accessible
carbon sources like sugar or starch. Others are specialized parasites.
Zygosporangia The name of the class is derived from
the way in which they reproduce sexually by the physical blending - fusion or
conjugation
- of morphologically similar gametangia
to form a zygosporangium (the teleomorphic phase).
'Zygos' is Greek
for a yoke or joining. The gametangia arise from hyphae of a single mycelium in
homothallic
species, or from different but sexually compatible mycelia in heterothallic
species. Zygosporangia usually develop thick walls, and act as resting spores.
Below, left, is a photograph of a petri dish containing nutritive agar
medium. Two sexually compatible strains of Phycomyces blakesleeanus
[There is a possibly apocryphal story that the author of
the species did not like Blakeslee] were
inoculated on opposite sides of the plate (large dark spots). The mycelia spread across the
agar surface. Where
they met, a line of zygosporangia with spiny appendages formed. The right-hand
picture shows what these developing zygosporangia look like under the dissecting microscope.


The four diagrams below show how a zygosporangium
of Phycomyces develops.

Note that there isn't any sexual differentiation in size or shape here:
since we can't call them male and female, we simply label the mycelia '+'
and '-'
When
compatible mycelia of Phycomyces blakesleeanus meet, individual hyphae establish
intimate contact, developing finger-like outgrowths and seeming to grapple
with one another. This lets them exchange chemical signals which
confirm that they are sexually compatible. Then the two hyphae grow
apart again, only to loop back, swelling as they approach each other, and finally meeting
head-on. They have become gametangia,
which fuse when their tips touch.
After the walls between the two gametangial tips have broken down and their multinucleate
contents have mixed, the mixture is quickly isolated by two septa, and the paired-off
nuclei fuse. The structure is now called a zygosporangium, and it
develops a thick and often ornamented wall, even while still supported on either side by
the former gametangia, which are now called suspensors.
Although the two suspensors are now just empty appendages, they make it easy to recognize
a zygosporangium when you see one. Here's one from Rhizopus.
Phylum 2 - Zygomycota -- conjugating Fungi
Class Zygomycetes
Orders Mucorales, Entomophthorales, Zoopagales, Kickxellales
Class Trichomycetes
Orders Harpellales, Amoebidiales, Asellariales, Eccrinales
Phylum 3 - Glomeromycota -- arbuscular mycorrhizal fungi
Class Glomeromycetes
New Phylogenetic System (more complex)
Phylum 4
Zygomycota
Subphylum Mucoromycotina
Order
Mucorales
Order
Endogonales
Order
Mortierellales
Groups currently of uncertain position
Subphylum
Entomophthoromycotina
Order
Entomophthorales
Subphylum
Zoopagomycotina
Order
Zoopagales
Subphylum
Kickxellomycotina
Order
Kickxellales
Order
Dimargaritales
Order
Harpellales
Order
Asellariales
Phylum 5 Glomeromycota
Class
Glomeromycetes
Phylum (?) Microsporidia
(e.g. Nosema,
intracellular parasites, mainly of insects, formerly considered protozoa,
but now regarded as extremely reduced fungi) see Corradi & Keeling
2009.
Phylum 4 Zygomycota
-- Conjugating Fungi

Anamorphs - You won't often see zygosporangia in
field collections, though I sometimes find a homothallic Syzygites producing them profusely
on wild mushrooms. But asexual or anamorphic
phases of zygomycetes are easy to find on mouldy bread or peaches, or on horse dung. A
number of examples are illustrated below.
Collect some fresh horse dung, keep it in a damp chamber, and look at it through a
dissecting microscope, or even a hand lens, every day. You should be able to follow
a sequence of specialized coprophilous
fungi - and the first to develop will probably be the spectacular anamorph of Pilobolus,
which is discussed below and in Chapter 11. The asexual mitospores are usually formed inside mitosporangia borne at the tips of
specialized sporangiophores. Zygomycete cell walls are mainly of chitin and the nuclei in their vegetative
hyphae are haploid. Now
for a taxonomic survey of the phylum and its two classes.
Traditional Class
Zygomycetes.
6 orders, 29 families, 120 genera, almost
800 species.
I will introduce you to four orders: Mucorales
- Entomophthorales - Zoopagales
- Kickxellales
Note that the last three are now considered to be
rather separate from the traditional Zygomycota, and are placed in
distinct Subphyla.
1) Order Mucorales 13 families, 56 genera, 300 species. This order includes all the common saprobic zygomycetes. Here belong the ubiquitous bread mould, Rhizopus stolonifer (below), and the equally common genus Mucor. This Order is now placed in the Subphylum Mucoromycotina
The classification outlined here has been
built on studies of morphology, development and ecology. But now
molecular biology is telling us that some of our assumptions are
incorrect. For example, the Mucoraceae is now believed to be
polyphyletic, as are the Thamnidiaceae, Chaetocladiaceae and Radiomycetaceae.
Not only that, but some of the genera, such as Mucor, Absidia and
Backusella are also polyphyletic, with their species distributed among
different clades. But since the new classification has not yet appeared
in a complete form, I will present a version of the extant classification, with
the warning that it will eventually undergo major revision.
The globose mitosporangia of the Mucoraceae are produced at the ends of
tall, stout, simple or branched hyphae called sporangiophores
(below, left). The sporangia often look dark when mature (below,
right). A
common name for this group is 'pin moulds.'


Each sporangium contains
hundreds of non-motile, asexual spores (SEM - below) within a delicate outer
membrane called a peridium.

The trade-mark of the family Mucoraceae, as recognized until very recently, is a swollen extension of the sporangiophore called a columella, which protrudes like a balloon into the sporangium (below).

The columella often persists after the delicate outer skin or peridium of the sporangium has disappeared and the sporangiospores have been dispersed. It persists in a collapsed condition in Rhizopus,
giving the former sporangia a rather puzzling, umbrella-like appearance


Spinellus is a parasitic member of the Mucoraceae which attacks agarics (see Chapter 5B) - especially species of Mycena.

Other families of the Mucorales often have fewer spores per sporangium, and their sporangia often have no columella.
Thamnidium elegans (Thamnidiaceae) (below), seems to compromise. Its tall sporangiophores have one large, terminal, columellate sporangium, but lower on the stalk there are branches which fork repeatedly in a dichotomous manner, the final branchlets ending in tiny mitosporangia (sporangioles) which contain only a few spores.


The reductionist tendency is also evident in Helicostylum (Thamnidiaceae) (above), which has 10-20 spores per sporangium, and in Blakeslea trispora (Choanephoraceae) (below), which has only three spores per sporangium.

This trend reaches its logical conclusion in Cunninghamella (Cunninghamellaceae), which has only one spore per mitosporangium. Now each of the many mitosporangia becomes detached and acts as a dispersal unit.
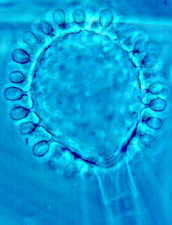

Although zygomycetes can go through cycle after cycle - spore > mycelium > sporangium > spore - producing only the anamorph, they sometimes form sexual zygosporangia (the teleomorph), perhaps as a survival mechanism, perhaps for the benefits of genetic recombination, or perhaps just because compatible strains have met. The photo (below) shows a developing zygosporangium (teleomorph) and a mature anamorph of Phycomyces.

The anamorph-teleomorph alternation is diagrammed below for one of the commonest and most successful members of the Mucorales, Rhizopus stolonifer. Note that when the zygosporangium germinates, it produces a mitosporangium.

Zygosporangia vary in minor ways from one genus to another, and among families and orders, but they are generally rather similar: if they are present, they are the easiest way to tell if a fungus is a zygomycete.
By contrast, the anamorphs of zygomycetes -- mitosporangia and the structures on which they are borne -- have evolved some amazing and bizarre adaptations. This contrast between teleomorphic constancy and anamorphic diversity is presumably the result of differing evolutionary pressures. Long-term survival, one of the main objectives of the teleomorph, is presumably best ensured by structures with minimal surface area and thick, protective walls. Dispersal, apparently the main purpose of anamorphs, can be achieved in many ways. Let's look at three of the more specialized zygomycetous anamorphs. Pilobolus crystallinus (Pilobolaceae, below) is an atypical but fascinating coprophilous (dung-inhabiting) member of the order Mucorales. It grows very rapidly, and is one of the first fungi to fruit in the extended succession that occurs on dung (see Chapter 11). Its unbranched sporangiophores are 2-4 cm tall, and have a unique explosive dispersal mechanism.


Beneath the black apical mitosporangium is a lens-like subsporangial vesicle,
with a light-sensitive 'retina' at its base that controls the growth of the sporangiophore
very precisely (above), aiming it accurately toward any light source. In a word, it is
phototropic.
Osmotically active compounds cause
pressure in the sporangiophore and the subsporangial vesicle to build up until it is more
than 100 pounds per square inch (7 kilograms per square centimetre). This eventually
causes the vesicle to explode, hurling the black sporangium away to a distance of up to 2
metres, directly toward the light. The mucilaginous contents of the subsporangial vesicle
go with the sporangium, and glue it to whatever it lands on.
There is a
high-speed movie of the shooting process on YouTube:
click here
Another YouTube piece shows similar footage, but adds a commentary which discusses the enormous acceleration imparted to the sporangium: click here
If that doesn't work for any reason, go to
YouTube and enter 'fungal cannon'.
This movie will surprise you, both by the acceleration involved, and by
the fact that the entire contents of the sporangium follow the spore mass
in a l;ong tail, almost certainly helping to glue it to a new (edible)
substrate.

This picture has the Pilobolus sporangia 'aiming' directly
into the camera lens, and the refraction of the subsporangial vesicle at
top right shows the yellow pigment in the 'retina'
Can you explain why Pilobolus
needs such a specialized mechanism for spore dispersal: such a powerful cannon, so
carefully aimed? You can find the answer in Chapter 11.
Note that the originality of Pilobolus extends only to the
behaviour of its anamorph - the teleomorph (the zygosporangium, shown here), is fairly
conventional.

This image, which looks completely out of place, is
actually of a highly original dance group that named itself Pilobolus.
You can catch them occasionally on television.

2) Order
Entomophthorales. As the name implies, these fungi often attack
insects. They are now placed in
Subphylum
Entomophthoromycotina

Entomophthora muscae infects, and eventually kills, houseflies.
Dying flies, their bodies riddled by the fungus, usually crawl into exposed situations --
I find them on windows, and on the growing tips of shrubs in my garden -- where the fungal
infection bursts through the insects' exoskeleton and produces tightly-packed masses of
sporangiophores (above and below).


Each sporangiophore bears one unicellular, sticky mitosporangium that is shot away at
maturity. When the fly dies on a window, this barrage produces a whitish halo of
mitosporangia on the glass. These sporangia can infect other unsuspecting flies that come
to pay their last respects. As you may already have guessed, species of Entomophthora
are being investigated for their potential in biological control of insect pests (see Chapter 14).
Note again that the zygosporangia of Entomophthora, though
developing in an unusual way, by the fusion of hyphal bodies inside the fly, are still
recognizable as zygosporangia.

3) Order Zoopagales
parasites of fungi, nematodes, amoebae, etc.
Many taxa produce merosporangia (sporangia that break up at maturity,
looking rather like the thallic-arthric conidia of some ascomycetes and
basidiomycetes). They are now placed in
Subphylum
Zoopagomycotina.
Piptocephalis (below) is a parasite of Mucorales, and occurs
commonly on dung. Note the merosporangia.


Syncephalis (below) is another parasite of Mucorales.
The photomicrograph shows a crown of young merosporangia arising
from a vesicle at the apex of a tall sporangiophore.

4) Order
Kickxellales
(Named after a mycologist called Kickx). These are now
placed in a distinct Subphylum
Kickxellomycotina, along with two orders
of the former Class Trichomycetes.
Most genera have only one or two species, and they are generally saprobes
isolated from faeces or soil (though there are three mycoparasites).
Members of this order are atypical of the
Zygomycetes in that they often have regularly septate hyphae
with septal plugs in the septal pores. Their teleomorphs are
unremarkable, but they develop some of the most complex anamorphs known. I
will show you one or two of these.

I have found Coemansia (above) on bat dung from a cave. Its tall
sporangiophore bears many fertile side branches called sporocladia. Each
of these produces a row of lateral cells called pseudophialides (true
phialides are discussed in Chapter 4). Finally, from the apex of each pseudophialide
arises an elongate, one-spored mitosporangium (a sporangiole).
That's complex enough, but it looks simple beside Spirodactylon
(below).
This, surely the most elaborate of all zygomycetous anamorphs, grows on the
dung of rats in Death Valley, California, and produces a tall,
dichotomously branched sporangiophore (a, b) that is repeatedly thrown into tight
coils (c) Within these coils
arise the sporocladia (d), which bear pseudophialides, that in turn bear one-spored
sporangioles.

It is hard to imagine why this strange configuration might have evolved, until one learns
that the fungus grows on mouse and rat dung. Coprophilous fungi have various highly
evolved strategies for getting back inside the gut of the animals that produce their
preferred substrate. This isn't too difficult for genera like Pilobolus, that
grow on herbivore dung, since all they have to do is get their spores onto the animal's
food, which is all around. But rats and mice are not herbivores, and it is essentially
impossible for the fungus to ensure that its spores will be present on their food. The
only alternative (as I see it) is to attach spores to the animal itself, in the hope that
they will be ingested during grooming activities. Rats and mice are creatures of habit,
using well-trodden paths each day. Along these trails they deposit dung, and there, later,
the coils of Spirodactylon become entangled in their hair. Only the zygosporangia
of the Kickxellales convince us that these strange fungi are indeed
related to the zygomycetes.
Traditional Class
Trichomycetes. 4 orders, 7 families, 52 genera, about 210 species.
Two groups formerly in this Class, Order
Harpellales and
Order Asellariales,
are now (2007) placed in the Subphylum
Kickxellomycotina, and the
other two orders, Amoebidiales and Eccrinales, are now seen to be
rather isolated, so the concept of the class has lost its integrity.
Trichomycetes are common in herbivorous or detritivorous arthropods, but rare in predaceous species. A single species of trichomycete can usually inhabit several related host species. They often seem to be commensals, doing little or no harm to their hosts: sometimes they may even be associated with faster growth of their hosts. But at least one species, Smittium morbosum, causes the death of mosquito larvae.
The Asellariales and Eccrinales are anamorphic orders, no zygospores having been seen in them.
These fungi are found attached to the gut lining of insects, crustacea and millipedes. M.M. White, a Canadian doing graduate studies at Kansas, has recently published the first report of a new genus of Harpellales in isopod Crustacea, of Trichomycetes in Trichoptera (Caddisflies), and of Eccrinales in Plecoptera (Stoneflies). These discoveries indicate that there is much to be learned about the taxonomy and host range of these groups.

This plate (above) of phase-contrast photomicrographs by Richard Benjamin (1979) shows the characteristic structures of some Trichomycetes. Top left (A) are developing trichospores of Smittium. Top right (B) are trichospores of Stachylina showing the hair-like appendages that give them their name. Bottom left (C) is a trichospore of Smittium. Bottom centre (D) is a developing zygosporangium of Trichozygospora, and bottom right (E) a released, mature zygosporangium with a collar and a bunch of hair-like appendages below it. Only this last structure (look at the conical "suspensors," top and bottom) places the Trichomycetes in phylum Zygomycota, since otherwise the group does not closely resemble the Zygomycetes.

Here are some illustrations (above) of Orphella catalaunica, a Trichomycete recently described from the hindgut of a stonefly nymph (Plecoptera, Insecta) in Spain.
Orphella is now placed with the Kickxellales
1 mature thallus
2, 3 sporulating heads with trichospores
4,5 holdfast cells with branches
6,7 sporulating heads
from Santamaria, S. and J. Girbal (1998) Mycol. Res. 102: 174-178.
Phylum 5 - Glomeromycota (arbuscular mycorrhizal fungi)
Class Glomeromycetes. 5 genera, about 170 species
The fifth eumycotan phylum is the very recently erected Glomeromycota. This new phylum (published in 2001) has relatively few members (fewer than 200 species have been described) but it is of enormous importance in the biosphere. There is a single class, the Glomeromycetes.
These
soil-inhabiting fungi were placed in the Zygomycota until very
recently, albeit rather tentatively, since they have never been seen to reproduce sexually. Nevertheless, they are extremely important, because their
hyphae enter the living root cells of perhaps 90% of all higher plants and establish with
them obligate mutualistic symbioses called arbuscular
mycorrhizae (AM) or endomycorrhizae. These
are discussed in detail in Chapter 17.
The large, thick-walled spores of the five currently recognized endomycorrhizal genera
are illustrated below (courtesy of Steve Bentivenga). Most mycologists
now prefer to merge a sixth genus, Sclerocystis, (which was
segregated on the basis of its many-spored fruiting structures) with Glomus.

Here are some thick-walled, lipid-filled spores of Glomus that
have been extracted from the soil by repeated sieving.

The photomicrographs below are of species of
Acaulospora with ornamented outer spore walls. It shows how
important microscopic features of these large spores are in the
classification of the group. The number and thickness of wall layers are
also important.
This plate is from Oehl et al. (2006).

AM fungi won't grow in axenic culture:
they must be associated with a plant root. Their generally very large and thick-walled
resting spores are common in most soils, and are stimulated to germinate by the proximity
of plant roots (almost any plant will do, because these fungi have such wide host-ranges).
Their usually non-septate hyphae spread through the soil and enter living roots, where
they develop structures that are diagnostic of the order: intracellular, finely
branched, tree-like arbuscules
(left) which are the interface across which the fungus exchanges mineral nutrients,
especially phosphorus (note the spelling!), for photosynthates (sugars, etc.) provided by the plant.

All Glomeromycetes produce arbuscules.
Many but not all species also develop lipid-filled structures
called vesicles or intramatrical spores inside plant roots, as this photomicrograph of a
root squash with Glomus versiforme shows.
This explains why the symbiosis has often, but incorrectly, been called
VAM, or vesicular-arbuscular mycorrhiza.

The soil-inhabiting mycelium is very efficient at mobilizing insoluble phosphorus and
translocating (moving) it to the plant. Since phosphorus is often the limiting nutrient
for plant growth, AM fungi help plants to thrive in poor soils. These fungi are
therefore vital in many natural habitats, and of great potential value in agriculture.
Again, for details consult Chapter 17.
Go to Chapter 4a
© Mycologue publications 2020
Further Reading
Here is the first good website all about Zygomycetes, with some excellent photomicrographs... http://www.zygomycetes.org/
Benjamin RK (1959) The merosporangiferous Mucorales. Aliso 4:321-453.
Benjamin RK (1979) Zygomycetes and their spores. pp 573-621 (in) The Whole Fungus. Vol. 2. (ed) Kendrick B. National Museums of Canada, Ottawa.
Bentivenga SP (1998) Ecology and evolution of arbuscular mycorrhizal fungi. McIlvainea 13: 30-39.
Brundrett M, Melville L, Peterson L (eds) (1994) Practical Methods in Mycorrhiza Research. Mycologue Publ., 8727 Lochside Dr.,Sidney, BC V8L 1M8, Canada (www.mycolog.com).
Cerda-Olmedo E, Lepson ED (eds) (1987) Phycomyces. Cold Spring Harbor, N.Y.
Cole GT, Samson RA (1979) Patterns of Development in Conidial Fungi. Pitman, London. 190 pp.
Corradi N, Keeling PJ (2009) Microsporidia: a journey through radical taxonomical revisions. Fungal Biology Reviews 23: 1-8.
Fuller MS (ed) (1978) Lower Fungi in the Laboratory. Dept. of Botany, Univ. of Georgia, Athens.
Ingold CT (1978) The Biology of Mucor and its Allies. Edward Arnold, London.
Kendrick B, Berch
SM (1985) Mycorrhizae: applications in agriculture and
forestry. pp. 109-152 (in) Comprehensive Biotechnology. Vol. 3. (Ed.) C.
Robinson. Pergamon, Oxford.
Kurihara Y, Degawa Y, Tokumasu S (2004) Two
novel kickxellalean fungi, Mycoemilia scoparia gen.sp nov. and
Ramicandelaber brevisporus sp. nov. Mycol. Res. 108: 1143-1152.
Lichtwardt RW (1986) The Trichomycetes. Springer-Verlag, New York.
Morton JB, Benny GL (1990) Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): a new order, Glomales, two new suborders, Glomineae and Gigasporineae, and two new families, Acaulosporaceae and Gigasporaceae, with an emendation of Glomaceae. Mycotaxon 37: 471-491.
O'Donnell KL (1979) Zygomycetes in Culture. Department of Botany, University of Georgia, Athens
Oehl F, Sykorova Z, Redecker D, Wiemken A, Sieverding E (2006) Acaulospora alpina, a new arbuscular mycorrrhizal fungal species characteristic for high mountainous and alpine regions of the Swiss Alps. Mycologia 98: 286-294.
Schuessler A, Schwarzott D, Walker C (2001) A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol. Res. 105: 1413-1421.
White MM (1999) Legerioides, a new genus of Harpellales in isopods and other Trichomycetes from New England, U.S.A. Mycologia 91: 1021-1030
Zycha H, Siepmann R, Linnemann G (1969) Mucorales. Eine Beschreibung aller Gattungen und Arten dieser Pilzgruppe. Cramer, Lehre.