
The Fifth Kingdom - Chapter 2b
SIMPLE FUNGI
Now for some organisms that are generally regarded as fungi. As I mentioned in chapter 1, what we call "fungi" share many morphological and behavioural similarities in their assimilative (feeding) phase, but they do not have a uniform genetic background. They have evolved from at least two ancestral lineages. Three small phyla -- Oomycota, Hyphochytriomycota, and Chytridiomycota -- can produce cells that swim by means of one or two very fine whip-like extensions called flagella (rather like the tails of sperms). Molecular evidence tells us that they are members of two different Kingdoms, the first two being Chromistan in origin, the last (despite its flagellate phase) Eumycotan. The other three main-line, largely terrestrial fungal phyla never have motile cells, and they -- the Zygomycota, Ascomycota and Basidiomycota -- make up the rest of Kingdom Eumycota.
Here is a diagrammatic
summary of a study of fungal relationships based on a study of 1551
ribosomal sequences (the largest number yet considered in a single study) by
Tehler, Little and Farris (2003)

The first four groups in the diagram are agreed
to be members of the Chytridiomycota by all taxonomists. No
surprises there. But to see the Blastocladiales linked to the
main groups of Zygomycota is a shock. Will this linkage hold
up? Only future studies will tell. For the time being, make
your own comparisons in this and the next chapter...
Before I introduce you to the three flagellate phyla, a few
words about flagella (singular: flagellum).
Flagella are very long, narrow organelles,
essentially contractile extensions of a cell, which have the ability to beat, or make
whip-like motions. This beating action moves the cell
about in water, giving it the ability to swim up chemical gradients (that is, to move from
a lower concentration of a substance toward a higher concentration), such as those which
lead toward a sexual partner or a suitable host organism.
The amazing thing about flagella is that wherever we find them among eukaryotic
organisms (even in members of different eukaryotic
Kingdoms), they have essentially the same fine structure: along the shaft of the flagellum
run 9 pairs of peripheral microtubules and 2 central microtubules -- the 9 + 2
pattern. Each microtubule is built from a protein called tubulin, its subunits
arranged in 13 vertical stacks (count them in the picture below) around a hollow
centre.
This picture of a transverse section of a flagellum (which is about
one-sixth (0.16) of a micron thick) would look more or less the same whether the flagellum came
from one of the simple fungi discussed here, or from a protozoan such as Paramoecium
(which has lots of short flagella called cilia all over the outside of the
cell), or from a unicellular or colonial green alga, a sea gooseberry
(phylum Ctenophora) which has plates of cilia that beat in rhythm and move the whole macroscopic
organism, or from the sperm (male gamete) of a brown alga, a sea urchin, a moss, a fern,
or
a human. You see, all eukaryotes really are related!

The somatic phases of these microscopic fungi vary widely in appearance, but when they undergo asexual reproduction most produce zoospores with a single, backwardly directed, whiplash flagellum. We'll look at members of several orders: the Chytridiales, Spizellomycetales, Blastocladiales, and Monoblepharidales. Although these orders were formerly separated on the morphology of their microscopic thalli, we now know that this is too variable to be reliable. They are now divided between two phyla.
Emphasis has switched to ultrastructural features of the zoospore, some of which are illustrated below. Unfortunately, although conservative, and therefore taxonomically valuable, these characters can be seen only in the transmission electron microscope after elaborate preparative techniques. Hyphochytrium is illustrated here (left-hand diagram) for comparison with the various chytridiomycetes.
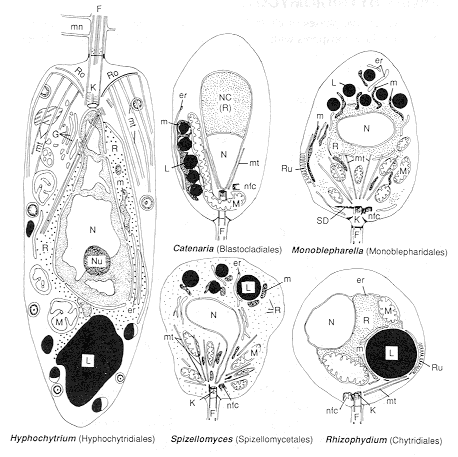
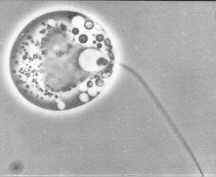
Fortunately, you can see enough of an Allomyces zoospore (above) under the light microscope (phase contrast) to identify the Phylum Blastocladiomycota.
Orders Chytridiales, Spizellomycetales and Rhizophydiales
These orders look very alike under the light microscope, and it takes an expert to tell them apart. However, most Chytridiales are aquatic, while most Spizellomycetales live in soil. Until important differences were found in the ultrastructure of their zoospores (above), those two orders were considered to be one. These simple fungi do not produce hyphae. They are often parasitic, and their assimilative thallus often consists of a single cell.
The thallus is either:
(1) entirely converted into a reproductive sporangium (the holocarpic mode, as in Olpidium brassicae (top right of diagram), or
(2) differentiated into assimilative rhizoids and a sporangium (the eucarpic mode), as in Spizellomyces punctatus (top left of diagram), or
Cladochytrium (bottom of diagram) which has a more extensive thallus...

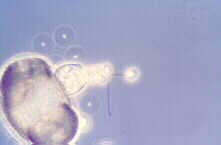
...or Rhizophydium (Rhizophydiales), seen here growing in culture:
this is a beautiful example of the eucarpic mode.

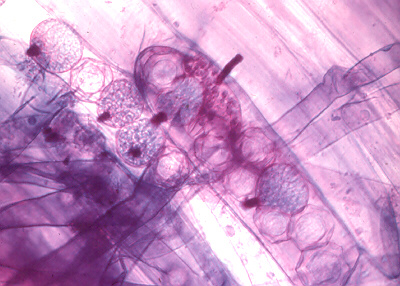
Other chytrids have a more extensive system of rhizoids, called rhizomycelium, which may nourish several sporangia, as in Cladochytrium (above). We describe this multisporangial condition as polycentric to differentiate it from the monocentric forms just mentioned, which produce only a single sporangium. The difference between rhizoids and rhizomycelium is that rhizoids generally have no nuclei, and are usually less than a millimetre long, while rhizomycelia contain nuclei and can be much more extensive. You might be interested in the activities of some of the members of these two orders, including the fungi just named. The holocarpic Olpidium brassicae (above) does not itself cause much damage to plant roots, but is the vector of some nasty plant viruses. The eucarpic Spizellomyces punctatus (above) and Chytridium lagenaria (below) parasitize pollen grains.
The four time-lapse photomicrographs below follow the release of the uniflagellate zoospores of Chytridium lagenaria after it has plundered a pine pollen grain (the large lobed structure at lower left in each picture).



At least one chytrid is a serious problem for farmers: Synchytrium endobioticum causes wart disease of potato. This fungus produces dark brown, cauliflower-like growths on the tubers (bottom), and a catastrophic reduction in yield. The pathogen is widespread in Europe, and has spread to Newfoundland, but resistant varieties of potato help to keep the disease under control.
However, in Fall 2000, a small corner of one field in Prince Edward Island, Canada, was found to be affected by the disease. This brought an immediate embargo on shipments from PEI to the USA, and as of January 2001, PEI growers stood to lose at least $36 million -- some of the 650 potato farmers in PEI may even lose their farms. Many seasonal employees also lost their jobs.

Other microscopic chytrids parasitize algae, and can be so numerous as to cause epidemics which significantly, if temporarily, reduce primary production in lakes.
The algal host here is Spirogyra, a green alga...

...while that shown here is Ceratium, a dinoflagellate.

The Frog Problem...
Most recently, a eucarpic chytrid has been found attacking amphibia in many part of the world. It has been associated with significant die-offs of frogs in Australia, Central America, and at the National Zoo in Washington, D.C., U.S.A. The condition has been quite reasonably called "chytridiomycosis" since no other organisms are yet known to be involved. The fungus lives inside epidermal cells and causes thickening of the skin which may interfere with normal respiration (frogs breathe partly through their skins). Its zoospores have ultrastructural characters (the kind that can be seen only under an electron microscope) that put it into a new genus, Batrachochytrium. Joyce Longcore, the chytrid taxonomist who determined this, published a short article in 'Inoculum', the newsletter of the Mycological Society of America for October 1998, and gave illustrations of both the eucarpic thallus and the zoospores. The paper describing the new genus appeared in Mycologia in April of 1999, and the full reference is given at the end of the chapter.


No one knows why this fungus would suddenly begin killing frogs in places as diverse as Australia and Panama. The fungus may have been transported to these places only recently - perhaps even on the boots or equipment of researchers studying the disappearance of frogs. Or the fungus may have been present for a long time but frogs are now succumbing because their immune systems have been impaired by recent environmental changes. One obvious change is increased ultraviolet light, which is known to damage immune systems of animals. Recently, chlorinated chemicals released by humans have attacked the ozone layer in the upper atmosphere, allowing 10% more U/V light to reach the Earth's surface. Industrial chemicals may also be damaging the frogs' immune systems. Retinoids are under suspicion because they cause birth defects in many animals, including frogs and humans. Accutane, used to treat acne, is a retinoid known to cause birth defects in humans. If you are interested in pursuing this topic, I suggest you search for more information on the internet: check out the archives of Rachel, an internet environmental magazine. In 2013 a new problem fungus has been discovered that attacks salamanders. It has been named Batrachochytrium salamandrivorans. Check out Martel et al. 2013 PNAS 110: 15325-15329.
Sexual reproduction in
Chytridiales and Spizellomycetales needs to be reexamined: it used
to be assumed that any zoospore with two flagella,
and every resting spore, resulted from nuclear fusion. Now we know that some biflagellate
zoospores originate by incomplete differentiation of cytoplasm during zoospore formation,
and that many resting spores are just thick-walled asexual sporangia which can survive dry
periods.
This picture shows resting spores of Olpidium
brassicae in wheat.
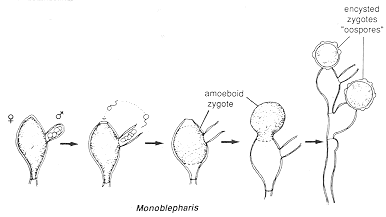
Sperms are often released before the adjacent oogonium is ripe. This may be a mechanism for avoiding self-fertilization, and ensuring outbreeding (called heterothallism in fungi). After the egg has been fertilized, the resulting zygote becomes amoeboid, moves out onto the top of the oogonium, and encysts, developing a thick wall (right-hand diagram). Meiosis probably occurs when this resting spore germinates, producing a germ tube (another name for a first hypha).

Some new and different chytridiomycete-like fungi!
Phylum 3 Neocallimastigomycota
Order Neocallimastigales Just to spoil our unitary picture of the chytridiomycetes, in 1975 Orpin discovered some new and very different fungi living in the rumens of large herbivorous mammals. These fungi, mostly species of Neocallimastix, were obligately anaerobic. They resembled their aerobic relatives in many ways, but had no mitochondria, and often had multiflagellate zoospores. So far, seventeen species of anaerobic chytrids have been described, representing five genera. They produce rhizomycelia which efficiently penetrate plant material, and have enzymes that break down cellulose even more effectively than the cellulases of Trichoderma. Molecular data now places these fungi in a distinct Phylum Neocallimastigomycota.
This drawing (after Barr 2001) shows the features of the zoospore of the Neocallimastigales. Note the roots of the multiple cilia (flagella) at the bottom of the drawing.

Although these microscopic fungi vary in so many things: in the morphology of their assimilative phase, in their patterns of sexuality, and in their adoption of parasitic or saprobic lifestyles; most of them have motile spores (zoospores or gametes), with one flagellum at the back (posteriorly uniflagellate - the cell swims like a sperm) [but note the often multiflagellate Callimastigales], and their cell walls, like those of the other, more complex, eumycotan fungi, are largely made of chitin (a polysaccharide very similar to the stuff of which insect exoskeletons are made). They synthesize lysine by the same pathway and the more advanced members of the group produce true hyphae. These fungi apparently represent modern survivals of the ancestral lines that evolved into the eumycotan fungi.
This group is like the chytridiomycetes in many ways: they live in fresh water or soil; they can be parasites or saprobes; they may be holocarpic or eucarpic, the latter kind having assimilative rhizoids; the sporangia release uniflagellate zoospores.
So why aren't they chytridiomycetes? A single character visible under the light microscope gave the original clue, and was soon supplemented by several ultrastructural features accessible only to the electron microscope (above). The 'visible' feature concerns the behaviour of the flagellum on the zoospore. In chytridiomycetes this propels the spore from the rear, but in hyphochytriomycetes the flagellum is seen to be attached at the front of the cell, and so it works like a swimmer's arm or an oar.
Other differences are revealed by the transmission electron microscope. In chytridiomycetes the axis of the flagellum is smooth (a whiplash flagellum), while in the hyphochytrids the axis of the flagellum bears many fine lateral filaments, called mastigonemes, that give it the name tinsel flagellum (the next illustration - the SEM picture below - shows both kinds of flagellum). These may appear to be insignificant differences, but biologists consider the numbers and kinds of flagella on zoospores to be an extremely conservative character -- one likely to remain unchanged over vast stretches of time, perhaps even hundreds of millions of years. This makes it an important indicator, which is amply confirmed by the ultrastructural differences illustrated above, and justifies the recognition of phylum Hyphochytriomycota. There are few well- documented species in this phylum, but Hyphochytrium catenoides is common in soil, and is often (like the chytrids, Spizellomyces and Chytridium) found in pollen when this is added to soil as 'bait' in the laboratory. This species may be of some importance in the natural biological control of plant pathogenic Oomycetes (see below), by parasitizing their oospores. No hyphochytrid has yet been seen to reproduce sexually.
Having just read about the minute but basic differences between the two previous groups, you won't be surprised to learn that similar inconspicuous features distinguish the oomycetes, our last and most important group of chromistan fungi. Although some authorities recognize 9 Orders (25 families, 95 genera and almost 700 species), we will treat only two Orders, Saprolegniales and Peronosporales (these words are hotlinked to the appropriate sections)
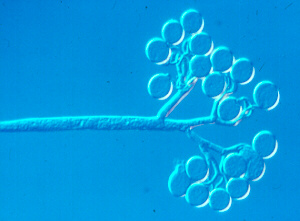
Basic features
(1) Oomycetous zoospores have two flagella: one tinsel, one whiplash (as shown clearly in the accompanying SEM), and these arise from the side of the cell, not the front or rear, as in other groups. The zoospores swim with their tinsel flagellum pointing forward, the whiplash flagellum directed backward. The tinsel consists of mastigonemes. Each of these has a long shaft with two divergent hairs of different lengths at its tip (the apical hairs are not visible here). This is a unique, highly conserved structure that is one of the basic features of the Kingdom Chromista.

(2) Unlike the nuclei of all true fungi, those in assimilative hyphae of oomycetes are diploid, and
(3) The walls of the hyphae usually contain a cellulose-like material (a poorly crystallized hexose polymer), though this makes up only a fraction of the cell wall, and chitin has also been found in some oomycetes. All of these characters separate them from the eumycotan fungi. But the feature that gives the group its name is its oogamous sexual reproduction, and the fact that each zygote develops into a thick-walled, persistent oospore (Note, however, that fungal oogamy is not unique to oomycetes - some chytridiomycetes such as Monoblepharis do it, too.)
Chytridiomycetes and
hyphochytriomycetes are often
extremely inconspicuous, as befits organisms that can devote a lifetime to exploiting a
single pollen grain. Oomycetes, though they also include some holocarpic and eucarpic
unicells, often produce extensive hyphal networks (mycelia). These fungi, covering a dead
fish like a whitish fur coat, or devastating crops such as grapes, hops, lettuce, cabbage,
radishes, potatoes and tobacco, have spawned a number of common names -- water
moulds and white rusts, downy mildews and damping
off. We'll take a look at some examples.
Order Saprolegniales
The water mould, Saprolegnia parasitica, attacks fish and their
eggs. A related fungus called Achlya can be isolated from water with hemp seed
bait (below).
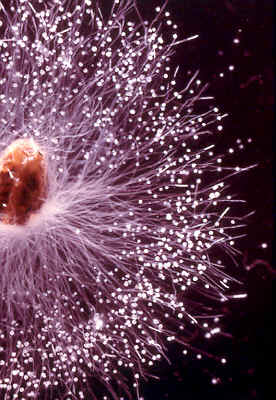
After establishing itself, the fungus reproduces sexually and asexually.
(1) The tips of the normally non-septate hyphae become modified into
long mitosporangia (you
can see them as short white radiating lines around the fringe of the colony.
Similar zoosporangia can be seen, more highly magnified, in the next two
illustrations below);
(2) many oogonia
(the numerous round white dots in the micrograph) also develop. These
are also seen at higher magnifications in the subsequent pair of pictures
(scroll down a little)


The asexual sporangia release biflagellate zoospores, often called swarm spores (below). These swim actively for a while, then encyst: they stop swimming and develop a thick wall.
Later, they germinate again as secondary zoospores which, if they are lucky, will find a new substrate and develop into new assimilative thalli. This encystment, followed by a repetition of germination, is a strategy that gives the spores a second chance at finding food if they aren't so lucky when first released.
Many Saprolegnia and Achlya species form compatible antheridia and oogonia on the same mycelium, which means that they are homothallic. Since the assimilative thallus is diploid, meiosis must take place inside the gametangia. Each globose oogonium contains several eggs, as you can see below.
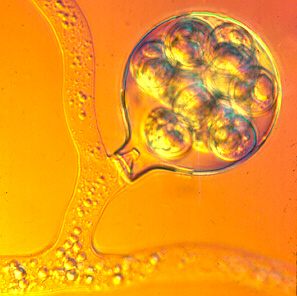

A number of antheridia may grow toward and touch a single oogonium, penetrating its wall at pre-formed thin spots and sending in fertilization tubes (above, right) which deliver the male nuclei to the eggs. Fertilization is more reliable because neither gamete is exposed to the vagaries of a free-swimming existence. The whole double life cycle is illustrated below.
The zygotes (oospores - lower far left) develop thick, resistant walls and obviously function as survival spores that can live through such catastrophes as the drying up of the pond or stream

Homothallic species have lost the enhanced variation provided by outbreeding, but they still benefit from the thick-walled resting oospores produced by the sexual cycle.
Order Peronosporales
Many members of this order are obligately parasitic on higher plants. In some cases
they cause epidemics that devastate important crops. The build-up of these epidemics is
made possible: (1) by our need to grow dense stands of single plant species (crops), and
(2) by aerial transmission of the fungi, which have evolved airborne mitosporangia (below,
left) These are often wrongly called conidia: they are analogous but not homologous
to those structures.
Oogonia are also formed, each containing a single egg (below, right). Sexual
reproduction is usually homothallic (the antheridium arises from the same thallus as the
oogonium, as you can see below, right).
We will examine representatives of five genera from three Families,
Pythiaceae (Pythium, Phytophthora), Peronosporaceae (Peronospora,
Plasmopara), and Albuginaceae (Albugo).

Damping-off disease of seedlings (Pythium - Pythiaceae)
Damping off of seedlings is a soil-borne disease, so its
causal agents, species of Pythium, have no need for airborne sporangia, since
they persist saprobically in most soils, and spread by zoospores during wet conditions.
When these motile cells find young plants, they cause infections which release toxins and
also produce a pectinase enzyme which dissolves the middle lamella that glues plant
cells
together.
Seedlings of many plants collapse rapidly when this disease strikes at the base
of their delicate shoots. Damping-off is, unhappily, familiar to gardeners who try to get
a head start on the growing season by germinating seeds indoors. The disease can be
controlled by using heat-sterilized soil, or by watering
seedlings with fungicides such as Zineb or No-damp.
The complete life cycle of Pythium
is illustrated here.

Late blight of potato (Phytophthora - Pythiaceae)
In 1991, the world produced 257 million tonnes of potatoes. In 2007
this had increased to almost 321 million tonnes. One reason is
that potatoes will grow in a wide range of climates. Another is
that potatoes yield from twice to four times as much as wheat or ice on
comparable areas. We in the West are particularly fond of them,
and find them incredibly versatile (to prove this to yourself, make a
list of all the ways in which potatoes can be prepared). Now the
rest of the world is catching on...
Potatoes were discovered (as far as the western world was concerned) by
the 16th century conquistadors when they
reached the Andes in South America. The
Spaniards saw the potential of this crop, but the potato was not well
received in much of Europe. This was perhaps because early
varieties were either bitter or bland. The French
naturalist, Raoul Combes, wrote "Here is the worst of all
vegetables." The famous French
gourmet, Brillat-Savarin, wrote "None for me...I know of nothing
more eminently tasteless." Early varieties also contained enough of
an alkaloid, solanine, to cause skin rashes which reminded people of the
dread disease, leprosy. Even in modern varieties,
there are significant quantities of solanine in green potatoes (make
sure you peel this green layer away before cooking).
In the mid-19th century,
a South American oomycete arrived in Europe and caused havoc. Unintentionally imported to Ireland
from the Andes, and reintroduced to a susceptible host from
which it had been separated 250 years earlier (when
potatoes were introduced to Europe), Phytophthora infestans
spread rapidly and wiped
out the Irish potato crop in the damp, cool summers of the years 1845-1847, causing
widespread famine in that poor, one-crop economy. At first, the disease was not attributed
to the fungus, but to an excess of water in the plants, or even to the effects of the
newly introduced steam locomotives.

However, the Reverend M.J. Berkeley, England's leading mycologist, drew
pictures of sporangiophores
emerging from the potato leaves [below], and correctly concluded that the fungus caused
the disease.
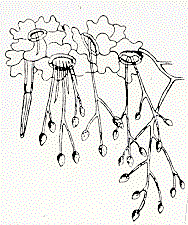
His contemporaries eventually admitted that he was right, and this
episode led to the founding of the discipline now known as Plant
Pathology.
Contemporary pictures from the London
Illustrated News (below, right) convey some of the misery caused by the recurring
epidemics. Ragged and starving peasant girls gleaned desperately in the fields for
anything edible (below, left), and whole families were forced to leave their homes forever.


Here is a contemporary report from one of the
worst affected areas: '...six famished and ghastly skeletons, to all
appearances dead, were huddled in a corner on some filthy straw...I
approached in horror, and found by a low moaning they were alive...Suffice
it to say, that in a few minutes, I was surrounded by at least 200 such
phantoms, such frightful spectres as no words can describe.'

Many died...
Many others sailed for North America. Some died during the voyage, and 187 doubly unfortunate souls were shipwrecked and
drowned off Forillon National Park near the tip of the Gaspé Peninsula, Quebec - in sight
of the promised land (below).


The
ravages of potato blight contributed to a million deaths, and drove
millions more to emigrate from Ireland. Ten years after the first
epidemic, the population of Ireland had crashed from 8 million to 4
million.
Until 1976 this heterothallic pathogen (except in its Central American homeland) was asexual, representing only one of the two mating types. In that dry year, many crop failures led to importation of potatoes that carried the other mating type with them. It has also been suggested that new genotypes were spread in seed tubers, in tomato tissues, and even in tropical storm systems. However it came about, new sexual populations have certainly supplanted the older asexual strain, and have led to a resurgence of the disease, which now costs the U.S. alone about $3 billion per year in attempts at control by spraying fungicides, by trying to breed resistant varieties of potato, etc.
In addition to its most infamous member, the genus Phytophthora contains about 60 other species. Some of them are also, as one might suspect, serious plant pathogens. Phytophthora sojae plagues soybean farmers in North America. Phytophthora megakarya attacks cacao trees in West Africa. A hybrid between P. cambivora and P. fragariae has killed about 10% of the Alder trees (Alnus spp.) in Britain and is now spreading to France, Holland, Sweden, Germany and Austria. P. cinnamomi is destroying Jarrah (Eucalyptus) forest and other natural ecosystems in western Australia, cork oak forests in Spain, and is a serious problem for growers of avocados, pineapples and ericaceous shrubs elsewhere in the world. A new disease that is killing thousands of oaks in California is caused by an unknown (undescribed) species of Phytophthora (see Forest Pathology section of chapter 12). No wonder the genus provides work for hundreds of plant pathologists and mycologists world-wide.
Downy mildew diseases (Plasmopara, Peronospora - Peronosporaceae)
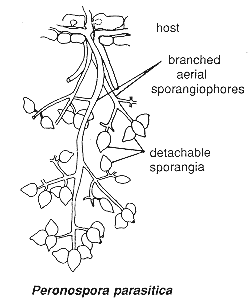
The downy mildews include blue mould of tobacco and downy mildew of grape (Potato blight is technically also a downy mildew). Since these diseases also have historic or economic importance, I'll tell you something about them. In this group the mitosporangia are borne on highly differentiated, branched, aerial sporangiophores, as shown below.
The sporangia don't just release zoospores but are themselves set free and blown or splashed away. The sporangia of Peronospora (below) germinate by producing a hypha, though those of most other members of the group still release motile zoospores.


Of course, this family also produces diagnostic oospores. A thick-walled, resistant oospore of Peronospora (growing on Chenopodium) is shown below (photo. courtesy of Jose Rodriguez)

Downy mildew of Grape (Plasmopara - Peronosporaceae)
Plasmopara viticola, an oomycete native to America, causes downy mildew of grapes. It can be found attacking wild grapes every summer. But because it evolved along with its North American hosts (e.g., Vitis riparia), a biological balance has been struck, and the wild Vitis species aren't seriously damaged.
The phase-contrast photomicrograph below shows a single, characteristically branched sporangiophore of Plasmopara with one sporangium still attached.
When this fungus was accidentally introduced to Europe in the 1870's, it was a different story. The cultivated French grape vines (Vitis vinifera) had no resistance to the new pathogen, and were quickly devastated.


The life cycle of Plasmopara is illustrated at below. Look at this set of diagrams carefully, and decide at which stage you think it would be most vulnerable to chemical attack (Here's a clue - the answer begins with z....).
Only the concoction of Bordeaux Mixture, one of the world's first practical fungicides, by a University Professor (yes, we Profs. do occasionally have good ideas!) saved the day. The rather strange story behind this invention is told in Chapter 13

This picture shows the bluish cast of grape leaves which have been sprayed with Bordeaux
Mixture (I took this picture on the famous Côte d'Or near Beaune in France)

This photograph shows a poster I saw in France for a modern version of Bordeaux Mixture
(brought to you by the makers of Aspirin)

Blue mould of Tobacco (Peronospora - Peronosporaceae)
My last example of a downy mildew fungus is Peronospora tabacina, which causes
blue
mould of tobacco. This disease was first recorded in Ontario in 1938, was
epidemic in 1945-1947, and had not been seen since 1966. But in 1979, seedlings infected
with the pathogen were imported to Canada from the U.S., escaping detection at the border.

The weather that year favoured the development and spread of the fungus, and a large-scale epidemic ensued. About 30% of the Ontario crop, worth $100 million, was lost. The picture above shows what the pathogen does to tobacco leaves.
Blue mould helped to put the Ontario tobacco industry on a slippery slope, and the decline in tobacco acreage is still continuing, though it is now driven by changing societal attitudes toward smoking.
Peronospora once again became a serious problem (a coffin nail?) for Ontario tobacco growers in 1997.
This picture shows a sporangiophore of Peronospora.
Note how its branching differs from that of Plasmopara (shown in
the fifth picture up).
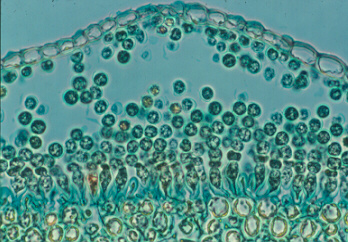
White rust disease of crucifers (Albugo - Albuginaceae)


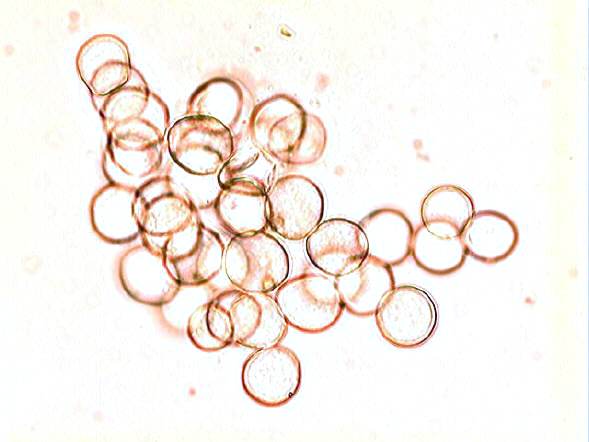
White rust of crucifers (cabbage, radish, etc. - all members of the family Brassicaceae) is caused by Albugo candida, which produces extensive, unmistakable white blisters on leaves and stems. The three images above are of a leaf and pods of 'honesty' (Lunaria annua) from our garden, and the spores they release.
These blisters contain innumerable unicellular mitosporangia
developing in chains from the tips of short, tightly packed sporangiophores (vertical
sections of blisters are shown in lower images). When the host epidermis bursts, the sporangia
are wind- or rainsplash-dispersed to other host plants, where each can germinate to
release eight biflagellate zoospores. Oogonia develop later, inside the host stem or
leaves, and sexual reproduction is usually heterothallic, or outbreeding.

Go to Chapter
3
Go to Table of Contents
© Mycologue Publications 2020
Further Reading and Web sites
Barr DJS (1990) Phylum Chytridiomycota. pp 454-466 (in) Handbook of Protoctista (eds) Margulis L, Corliss JO, Melkonian M, Chapman DJ. Jones and Bartlett, Boston.
Buczacki ST (ed) (1983) Zoosporic Plant Pathogens, a Modern Perspective. Academic Press, New York.
Dick MW (1990) Phylum Oomycota. pp 661-685 (in) Handbook of Protoctista
(eds) Margulis L, Corliss
JO, Melkonian M, Chapman
DJ.
Jones and Bartlett,
Boston.
Erwin DC, Ribiero OK (1996) Phytophthora diseases worldwide. 592 pp. APS Press, St. Paul.
Fuller MS (1990) Phylum Hyphochytriomycota. pp 380-387 (in) Handbook of Protoctista (Eds.) L. Margulis, J.O. Corliss, M. Melkonian and D.J. Chapman. Jones and Bartlett, Boston.
Fuller MS, Jaworski A (eds) (1987) Zoosporic Fungi in Teaching and Research. Southeastern Publishing Co., Athens.
Karling, J.S. (1977) Chytridiomycetarum Iconographia. Cramer, Vaduz.
Large EC (1962) The Advance of the Fungi. Dover, New York.
Longcore JE, Pessier
AP, Nichols DK (1999) Batrachochytrium dendrobatidis
gen. et sp. nov., a chytrid pathogenic to amphibians.
Mycologia 91:
219-227.
Margulis L, Corliss JO, Melkonian M, Chapman DJ (eds) (1990) Handbook of Protoctista. Jones and Bartlett, Boston.
Moore RT (2000) Mycological dispatches
[about Phytophthora].
Mycologist 14:93
Sparrow FK (1960) Aquatic Phycomycetes. 2nd Edn.
University of
Michigan Press, Ann Arbor.
Spencer DM (ed) (1981) The Downy Mildews. Academic Press, New York.
Tehler A,
Little DP, Farris JS (2003) The full-length phylogenetic tree from 1551
ribosomal sequences of chitinous fungi, Fungi.
Mycological Research 107:
901-916.